演習に関する学術用語の解説
(有用水産動植物)

竹原ステーションで実施される演習に関する学術用語の解説(他大学・広大の学部生向け)を 「共生」
「魚病など」
「プランクトン」
「海洋環境」
「無脊椎動物」
「有用水産動植物」
「海藻」
「魚類」
に纏めました。予習、復習に活用ください。
詳細は、各用語をクリックしてください。表中の図は、クリックで拡大されます。
| 有用水産動植物 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 遺伝子 | ゲノム | 減数分裂 | 二価染色体 | キアズマ | 受精 | 異数性 | |||||||
| アリル | メンデルの法則 | ホモ接合体・ヘテロ接合体 | 亜種 | 品種 | |||||||||
| 系統育種 | 遺伝的多様性 | 遺伝的浮動 | 創始者効果 | ビン首効果 | 近交弱勢 | ||||||||
| 染色体 | セントロメア | 動原体 | テロメア | (姉妹) 染色分体 | 相同染色体 | 核型 | |||||||
| 動原体型・腕比 | 倍数性 | 染色体操作 | 選抜育種 | 遺伝マーカー | 遺伝子座 | マイクロサテライトDNA | |||||||
| 顕性 (優性) | 潜性 (劣性) | 遺伝 (子) 型 | 表現型 | 形質 | ハーディー=ワインベルグの法則 | 有効集団サイズ | |||||||
| 人工種苗 | アコヤガイ | アマノリ | アオサ・アオノリ類 | キチン・キトサン | バイオミミクリー | 採苗 | |||||||
| 三倍体 | 極体 | 付着生物 | マボヤ | 生理活性物質 | 八方サンゴ類 | テトロドトキシン | |||||||
| テングサ | ミオグロビン | カロテノイド | K値 | ||||||||||
| ○引用文献 ・藻類ハンドブック 渡邉信ら編著 NTS 2012年 ・日本動物大百科 第7巻無脊椎動物 日高敏隆(監修)(1997)平凡社 ・三省堂 大辞林 http://www.weblio.jp/cat/dictionary/ssdjj(2017年6月閲覧) ・食の医学館 本多京子(2002) 小学館 ・日本かきセンターHP http://www.oyster-center.com/oyster/(2017年5月閲覧) ・デジタル大辞泉 小学館 http://www.daijisen.jp/(2017年5月閲覧) ・水圏生物科学入門 会田 勝美(編集)恒星社厚生閣 2009年 ・水産海洋ハンドブック第3版 竹内俊郎他(2016)生物研究社 ・荒井克俊 (1997) 水産増養殖における染色体操作の現状. 水産増殖, 45: 411–416. ・荒井克俊 (2017) 第6章 染色体操作と育種. 中嶋正道, 荒井克俊, 岡本信明, 谷口順彦 (編)「水産遺伝育種学」.東北大学出版会, 仙台, 99–118. ・高井明徳 (2016) 動物の染色体と核型. 人と環境, 9: 1–16. ・谷口順彦 (2007) 魚類集団の遺伝的多様性の保全と利用に関する研究. 日本水産学会誌, 73: 408–420. ・日本遺伝学会 (監修編) (2017) 遺伝単. NTS. 東京. pp371. |
|||||||||||||
有用水産動植物
遺伝子 gene遺伝形質を規定する因子。1900年代初頭まで仮想的な扱いであったが、その後ゲノム (「ゲノム」参照)、あるいは染色体の特定の場所 (遺伝子座位;「遺伝子座位」参照) に存在することが明らかとなった。20世紀中盤には、核酸が遺伝子の実体であることが証明された。遺伝子は、ゲノムDNAあるいはRNA上の特定の領域に存在し、塩基配列によってタンパク質をコードしている。タンパク質の1次構造であるアミノ酸配列をコードする遺伝子は構造遺伝子と呼ばれる。その他にも遺伝子の発現調節を行う調節遺伝子、核ゲノム以外にもミトコンドリア (細胞小器官) 内に存在するミトコンドリア遺伝子などが存在する。 |
||
ゲノム genome1920年にドイツの植物学者ハウス・ウィンクラー (H. Winkler) によって造られた造語で、遺伝子 geneと染色体 chromosomeを掛け合わせた言葉。半数性の1組の染色体が持つDNA―遺伝情報の総体。ゲノム中にはタンパク質構成情報を含む「コード領域」と、それ以外の「非コード領域」が存在する。ゲノムの大きさ (ゲノムサイズ genome sizeあるいはC値 C-value) は生物間で大きく異なる。また、ミトコンドリアや葉緑体のような細胞小器官がもつゲノムと区別して、核がもつゲノムを核ゲノム、ミトコンドリアおよび葉緑体にそれぞれ含まれるゲノムをミトコンドリアゲノム、葉緑体ゲノムと呼ぶことも多い。 |
||
減数分裂 meiosis真核生物の配偶子形成時に起こる細胞分裂の様式の一つ。アリル (「アリル」参照) の組み合わせを多様化することにより、遺伝的多様化を生み出す主なプロセス。染色体数2n=6の生物の場合、形成される配偶子 (卵もしくは精子) 染色体の組み合わせは23=8通りとなり、配偶子同士の組み合わせ、つまり次世代における染色体の組み合わせは82=64通りとなる。ヒトの場合、染色体数は2n=36であるので、配偶子染色体の組み合わせは223=8,388,608通りとなり、2個体間から生まれる次世代では8,388,6082、約70兆の組み合わせが生まれる。減数分裂は大きく第一減数分裂と第二減数分裂に分けられる。第一減数分裂前期から中期にかけて、相同染色体は対合し二価染色体 (「相同染色体」「二価染色体」参照) が形成される。この時キアズマ形成に伴う乗換え (「キアズマ」参照) により、隣接するDNA分子との部分交換が起こる。この乗換えの重要性は、第二減数分裂時に配偶子に分配される染色体の多様性を高める点である。第一減数分裂に続いて起こる第二減数分裂は、DNAの複製を伴わない細胞分裂で、染色分体が娘細胞 (配偶子) に分離する。この時の分離・分配によって、上記の組み合わせが生じる。 一般的に発生の実験に使用されるウニ類 (棘皮動物) は、減数分裂を完了した状態で成熟卵が体外に放出される。しかしながら、多くの貝類 (軟体動物) の卵は、第一減数分裂前期もしくは中期で最終成熟に達し放卵され、受精可能となる。精子侵入後に減数分裂が再開され、第一極体、続いて第二極体 (「極体」参照) の放出が起こり減数分裂を完了する (以下「受精」参照)。また、魚類、両生類、哺乳類などの多くの脊椎動物では、第二減数分裂中期で分裂が停止しており、精子の侵入により減数分裂が再開される。ウニと同じ棘皮動物であるヒトデ類では、第一減数分裂中期に放卵される。 |
||
二価染色体 bivalent chromosome第一減数分裂前期から中期にかけて、相同染色体が互いに対合した染色体像。通常少なくとも1個のキアズマが観察される。 |
||
キアズマ chiasma第一減数分裂前期の接合期に、相同染色体の非姉妹染色分体同士が交差してX字型になった箇所。キアズマの部分で相同染色体間の遺伝的乗換え (crossing over)、すなわち相同染色体間の非姉妹染色分体での一部相互交換が生じたとされる。 |
||
受精 fertilizatione精子が卵に侵入し雄性前核と雌性前核の核融合が起こること。魚類をはじめとする多くの脊椎動物では、第二減数分裂中期で分裂が停止しており、卵内への精子侵入により減数分裂が再開される。精子侵入後、卵は第二極体を放出し第二減数分裂を完了する。減数分裂の完了により、卵核は雌性前核 (female pronucleus) となり、卵細胞内にとどまっている精子核由来の雄性前核 (male pronucleus) と融合することにより受精が完了する。一方、水産養殖・種苗生産現場において、受精卵を得る (受精させる) ために採卵した卵に精子を加える行為を授精あるいは媒精 (いずれもinsemination) と呼ぶ。 |
||
異数性 aneuploidy染色体数異常の一つ。一倍性染色体セット (ゲノム) から1~数個の染色体が過剰にある、あるいは失われた状態のこと。また、このような状態の個体、あるいは細胞のことを異数体 (aneuploidy) という。染色体数異常には他に倍数性 (「倍数性」参照) が挙げられるが、倍数体は一倍性染色体セットが三つ以上存在する(e.g. 全ての染色体が3本ずつある=三倍体)状態を指し、一倍性染色体セットから数個の染色体が過剰に存在 (e.g. 21番染色体のみ3本)、あるいは失われた状態である異数性と異なる。二倍体生物における異数性の例として、特定の相同染色体が3本 (すなわち1本増加) ある状態をトリソミー (trisomy)、1本不足している状態をモノソミー (monosomy) などが挙げられる。このような異数性は、染色体不分離 (chromosome non-disjunction)、つまり細胞分裂時に染色体が均等に分離されずに、2つの娘細胞の片方のみに染色体が移動することによって生じる。染色体不分離は体細胞分裂、減数分裂 (「減数分裂」参照) いずれでも起こるが、減数分裂時や発生初期での染色体不分離は個体全体に大きな影響を及ぼす。 |
||
アリル (アレル、対立遺伝子) alleleゲノム上の座位 (「遺伝子座位」参照) に注目した場合、その領域におけるDNA配列の変異によって生じる個々の遺伝子 (あるいは、特定のDNA分子) の型を指す言葉。二倍体生物の場合、1遺伝子座位に同一のアリルが存在する場合をホモ接合、異なる場合をヘテロ接合と呼ぶ。alleleに対し、伝統的に「対立遺伝子」という訳が当てられてきたが、alleleは上述したように遺伝子に限定されず、実際にSingle Nucleotide Polymorphism (SNP) やマイクロサテライト (「マイクロサテライトDNA」参照) などもalleleの1つとして扱う。そのため、近年では「アリル」あるいは「アレル」とカタカナで表記するのが一般的。アリルは生物の遺伝的多様性を理解する上で基本的かつ重要な要素の1つ。 もとの意味は「もう一つ別の」「同列であるが異なる」というギリシャ語に由来する。 |
||
メンデルの法則 Mendel's laws日本では「分離の法則」「顕性 (優性) の法則」「独立の法則」の3法則として扱われる。分離の法則 (メンデルの第1法則): ある遺伝子座位においてヘテロ接合の個体が持つ2種類のアリル“Aa”が、減数分裂の際に均等に配偶子に分離する。しばしば、“3:1”などの分離比の意味と誤解されることがあるため注意が必要。 顕性の法則 (メンデルの第2法則): “AA×aa”のように、それぞれ異なるアリルからなるホモ接合の個体を交配した場合、得られる雑種第1代の表現型は均一であり、顕性アリルの表現型を示す。 独立の法則 (メンデルの第3法則): 2つ以上の遺伝子座位に注目した場合、それぞれの座位のアリルは独立に伝達される。しかし、本法則は注目する遺伝子座位が別の染色体上、あるいは同一染色体上であってもそれらが遠く離れている場合に限って、すなわち連鎖関係にない場合に限り成り立つ。 |
||
ホモ接合体・ヘテロ接合体 homozygous・heterozygous二倍体生物の場合、1遺伝子座位に“AA”、あるいは “aa”のように同一のアリルが存在する場合をホモ接合、“Aa”のように異なる場合をヘテロ接合と呼ぶ。ヘテロ接合で表現型がいずれか1つのアリルに依存する (ホモ接合の表現型に一致する) 場合、そのアリルを顕性 (優性)、表現型として現れなかったもう一方のアリルを潜性 (劣性) と呼ぶ (「顕性 (優性)・潜性 (劣性)」参照)。 |
||
亜種 subspecies生物の分類体系において、種の下に置かれる階層。同種内で互いにそれぞれ固有の特徴を有する集団。亜種間では交配が可能 (それらは同種) であるため、亜種は有用形質の供給源として育種に使用されることがある。 |
||
品種 breed栽培生物の実用的形質に関して、他の集団とは区別し得る遺伝的性質を持った集団。 |
||
系統育種 line breedingある有用形質を保有する原種とその形質を付与したい栽培生物をそれぞれの系統とし、それらの雑種に対し、戻し交配と選択を繰り返し行うことで、既存の品種に新たな形質を付与し、新たな系統を作出すること。系統とは祖先が共通であり、遺伝子型 (「遺伝子型」参照) がほぼ等しい集団を指す。 |
||
遺伝的多様性 genetic variation集団内におけるアリル (「アリル」参照) の多様性のこと。自然災害や人間活動による集団の分断や個体数の減少は、集団の縮小とそれに伴う遺伝的多様性の減少を引き起こす (「遺伝的浮動」参照)。遺伝的多様性減少は、集団内での有害遺伝子の拡大・固定、環境変動への適応性の喪失や近交弱勢を引き起こすリスクを高め、最終的には集団の衰退・崩壊につながる。そのため、これまで、そしてこれからの水産資源管理・持続可能な利用のためにマイクロサテライトDNAなどの遺伝マーカー (「マイクロサテライトDNA」「遺伝マーカー」参照) を用いた野生集団の遺伝的多様性評価、さらには乱獲など人類の諸活動が対象種の集団構造に与える影響について評価が行われてきた。一方、系統や品種作出 (「品種」参照) においては、望ましい形質以外の対立遺伝子を集団内から除外することを目的としており、遺伝子型がほぼ等しい集団が作り出される (「選抜育種」「系統育種」参照)。しかし、品種作出の過程で目的形質以外の遺伝子座でも遺伝的多様性の低下 (均一化される) が生じる。そのため、野生集団と比較して、栽培品種は冷害などの環境悪化や病害虫の発生で深刻な被害を受けることがある。 |
||
遺伝的浮動 genetic drift偶然性によってアリル頻度の変化が起こり、特定のアリルが集団内に広がる現象。このアリル頻度の変化に自然選択は含まれない。特に遺伝的浮動は、集団サイズが小さい、あるいは何らかの要因によって集団縮小が起こった際に作用する (「遺伝的多様性」参照)。ハーディー=ワインベルグの平衡 (「ハーディー=ワインベルグの法則」参照) が成り立っていれば、遺伝的集団構造を変化させずにその集団は存続し続ける。しかしながら、ハーディー=ワインベルグの法則は、無限集団を前提としている。現実にはその集団がどんなに多数の個体で構成されていたとしても、有効集団サイズ (「有効集団サイズ」参照) は、有限であるためアリル頻度は遺伝的浮動によって変動する。代表的なものとして、創始者効果 (founder effect) とビン首効果 (bottle neck effect) が挙げられる。 |
||
創始者効果 founder effect少数の個体 (後の創始者となる) が元の集団から別れた後、新たな地域に侵入、繁殖することで、隔離された新個体群を形成した場合において、遺伝的浮動の影響により元集団と異なるアリル頻度の個体群が形成されること。 |
||
ビン首効果 bottle neck effect集団遺伝学上の概念であり、集団の大きさが一時的に激減することにより、遺伝的浮動の効果が大きくなり、進化的に中立なアリルが固定、あるいは消失しやすくなり、集団内の遺伝的均一化が促進されること。 |
||
近交弱勢 inbreeding depression近親交配を長期間続けることにより、大きさ、耐病性、多産性などの有用形質が低下する現象。近交弱勢が生じる原因として、アリル数の減少とホモ接合の遺伝子座が増加することで、潜性有害遺伝子の影響が出ることが原因と考えられている。 |
||
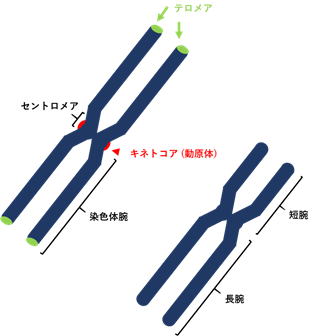
染色体 chromosome真核生物の細胞分裂の際 (分裂期; M期)、核内のDNA (クロマチン繊維) が凝集してできた棒状の構造体。染色体がその形態と機能を維持するためには、染色体の均等分配に係わるセントロメア、末端に位置し染色体の安定化にかかわるテロメア、DNA複製開始点の3つの要素が必要。セントロメアを挟んだ両側を染色体腕と言い、短い方、および長い方の腕をそれぞれ、短腕 (核型記載の記号として“p”が用いられる)、および長腕 (q) と呼ぶ。また、セントロメアの位置 (短腕と長腕の相対的長さの比率) に基づいて5つの動原体型に分けられる。 |
 |
|
セントロメア centromere染色分体を連結させている染色体の“くびれ (狭窄)”のように見える領域。紡錘糸が連結する (動原体が形成される) 領域でもあり、細胞分裂期における染色体の安定した分配制御に必須の染色体領域。 |
||
動原体 (キネトコア) kinetochoreセントロメア領域にある紡錘体が結合する特定構造。「動原体」という呼称について、これまでセントロメアとキネトコアを明確に区別せず用いてきたが、最近キネトコアを指す言葉として定着している。しかし、“Metacentric (中部動原体)”等の合成語の訳については、動原体が用いられる。多くの生物でキネトコアはセントロメア1カ所に局在しているが、染色体全体にわたってキネトコアが散在する「分散型動原体」を有す染色体 (全動原体染色体) がある。この染色体はくびれがなく、動物では線虫、チョウやガ、カメムシの仲間、植物ではカヤツリグサ、モウセンゴケなどで認められる。 |
||
テロメアtelomere染色体両腕の末端部分。染色体の形態を保持するために必須な領域。また、テロメアは細胞分裂に伴って起こるDNA末端の短縮が、遺伝子領域にまで及ぶのを防ぐ役割も持つ。細胞分裂に伴うテロメアの短縮が、一定の長さにまで達すると細胞分裂は停止し、このことを“ヘイフリック限界Hayflick limit”と呼ぶ。テロメア配列は5-8塩基からなる反復配列で構成されており、1978年に単細胞真核生物の繊毛虫の仲間Tetrahymena thermophilaで初めて明らかにされた [(TTAGGG)nタイプ]。海綿動物、軟体動物、棘皮動物、脊椎動物をはじめ動物界で広く確認されているのは (TTAGGG)nタイプであるが、節足動物 [(TTAGG)n]や線形動物 [(TTAGGC)n] といった分類群では、反復モチーフに違いがあることが知られている。 |
||
(姉妹) 染色分体分裂期に現れるセントロメアで結合された2本の染色体。細胞周期の複製期に1本の染色体から複製されたコピーの染色体。父親 (精子) と母親 (卵子) にそれぞれ由来する1対2本の同一の染色体を指す相同染色体と混同されることが多い。 |
||
相同染色体 homologous chromosome二倍体細胞内に存在する、父親 (精子) と母親 (卵子) にそれぞれ由来する1対2本の同一の染色体。第一次減数分裂時に互いに対合し、二価染色体を形成する。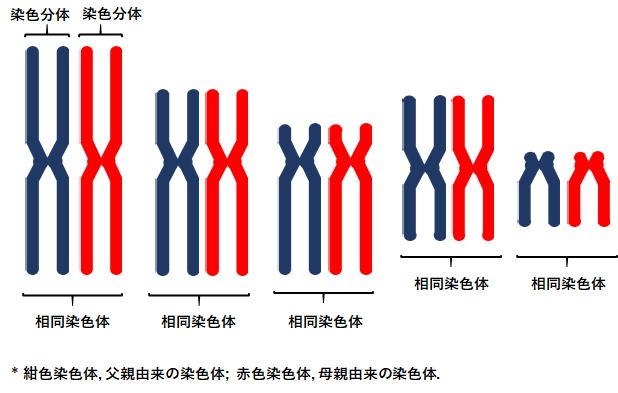 |
||
核型 Karyotype染色体の数、サイズ、バンドパターン、および動原体型等の形態的特徴に基づく染色体構成、または、それらを表した図や数値、記号で表したもの。 |
||
動原体型・腕比 Arm ratioセントロメアの位置、すなわち短腕と長腕の相対的長さの比率 (腕比) に基づく染色体の区分で、Levan et al. (1964)によるに形態分類が広く用いられている。セントロメアが中央付近にある染色体を中部動原体型染色体 Metacentric chromosome、(M; 1.0-1.7)、セントロメアが中央よりやや端に寄っているものを次中部動原体型染色体 Submetacentric chromosome (SM; 1.7-3.0)、セントロメアが端に近いものを次端部動原体型染色体 Subtelocentric chromosome (ST; 3.0-7.0)、セントロメアが端にあるものを端部動原体型染色体Acrocentric chromosome (A; 7.0-∞)、そしてセントロメアが末端部にあるものを末端部動原体型染色体Telocentric chromosome (T; 短腕なし)という。括弧内のアルファベットおよびアラビア数字は、それぞれ核型記載の記号および腕比 (長腕/短腕) を表す。 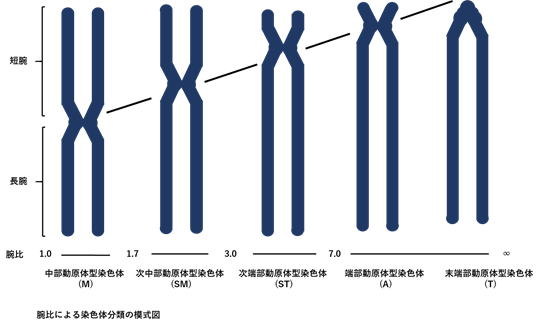 |
||
倍数性 polyploidy一倍性染色体セット (ゲノム) が三つ以上存在する状態。このような個体あるいは細胞を倍数体 (polyploid) と呼ぶ。同種由来の相同な染色体のみを持つ倍数体を同質倍数体 (autopolyploid)、異種由来の非相同な染色体をもつ雑種倍数体を異質倍数体 (allopolyploid)と呼ぶ。同質倍数体について、植物では倍数性の上昇に伴い、植物体が大型化するギガス性 (gigantism) が見られ、その可能性はマガキなどの貝類でも指摘されている。三倍体では、生殖腺の発達が抑制されることにより、可食部の比率が高くなる。また、性成熟は肉色、肉成分、食味などにも影響することから、不妊三倍体では肉質の改善が期待されている。広島県における同質三倍体牡蠣の「カキ小町」や長野県の全雌異質三倍体「信州サーモン」(四倍体ニジマス雌×二倍体ブラウントラウト雄) などが養殖生産されている。 |
||
染色体操作染色体セットの数と組み合わせに細胞分裂 (極体放出および卵割) の阻止や配隅子の遺伝的不活性化等の人為的な変更を加える技術。細胞分裂阻止には、中心体、紡錘体などの分裂装置 (微小管) 破壊による染色体移動の阻害 (核分裂阻止) と、収縮環のくびれ阻害による細胞分裂阻止がある。これら細胞分裂阻止による倍数体作出法として、低・高温による温度処理、高圧処理といった物理的処理、およびカフェイン、サイトカラシンBによる化学的処理がある。また、染色体操作は魚類の性操作法や不妊化法としても利用可能なことから、育種のみならず生殖統御技術としても体系化されている。 |
||
選抜 (選択) 育種 selective breedingある特定の有用形質をもつ個体もしくは集団を選びだし (=選抜し)、それらの中から継代的に選抜を繰り返し行うことで、形質の改良を目指す育種法の1つ。選抜育種の中には、一定の遺伝的特性を備える家系に由来する個体を選抜する家系選択法 (family selection) と、一定の有用形質を持つ個体に注目する個体選択法 (mass selection) がある。選抜育種の例として、欧州で古くから行われてきた、体表面にほとんど鱗を持たない突然変異を選抜したドイツゴイ、および米国で約30年に渡り行われてきた、高成長を指標にしたニジマスの選抜 (ドナルドソンニジマス) などが有名。 |
||
遺伝マーカー genetic marker遺伝的性質、あるいは親、祖先からの由来 (個人の特定、親子鑑定、および血統・品種) の検出を目的として作られたゲノムの特定領域。タンパク多型 (例: アイソザイム) やDNA多型 (例: マイクロサテライトDNA、一塩基多型 “SNP”) が遺伝的多様性評価のマーカーとして使用され、その多型性の程度が高ければマーカーとしての性能 (感度) が高いとされる。 |
||
遺伝子座 (位) locus特定の塩基や遺伝マーカー、配列などの染色体上での位置。商業ビルを染色体、そこに入っているショップをアリル (遺伝子や遺伝マーカー) に例えた場合、ショップが入っている場所が遺伝子座 (位) にあたる。 |
||
マイクロサテライトDNA microsatellite DNAゲノム中に散在する2-5塩基からなる短い反復配列。繰り返し回数に多型性があり、また親から子にこの繰り返し回数が遺伝することから、配列長を利用した遺伝マーカ―として利用される。生物的機能を担う部分であるアイソザイムと比較して、極めて高い多型性を持つことから、遺伝的多様性の評価や親子判別のツールとして使用される。 |
||
顕性 (優性) dominantその表現型が単一遺伝子に依存する場合、遺伝 (子) 型 (アリルの組み合わせ) が、ヘテロ接合の時でもホモ接合の時同様にその形質が現れること。従来の優性に代わる用語。ヒトのABO血液型では、A型B型の間に顕性・潜性の関係はなく、ヘテロ接合体で2つのアリルの形質が共に表現型として顕れることを共顕性codominantという。 |
||
潜性 (劣性) recessiveその表現型が単一遺伝子に依存する場合、遺伝 (子) 型 (アリルの組み合わせ) が、ヘテロ接合の時は現れず、ホモ接合の時にのみ、その形質が現れること。従来の劣性に代わる用語。 |
||
遺伝 (子) 型 genotype個体および生物集団について、特定の遺伝子座 (位) におけるアリルの組み合わせ。形質と関連する遺伝子座 (位) の場合、外面に現れた形質タイプのことを表現型という。表現型が同じであっても、遺伝子型が異なる場合もある。例えば顕性アリルをホモ接合で持つ赤花 (遺伝子型: AA) と潜性アリルの白花 (遺伝子型: aa) を交配させた場合、得られる雑種第一代の遺伝子型はAaとなり、AAの遺伝子型と異なるが、同一の表現型 (赤花) を示す。 |
||
表現型 phenotypeある遺伝 (子) 型に対して外面に現れた形質タイプのこと。 |
||
形質 trait生物の形態や機能上の特徴や性質。表現型として現れてくる要素的な遺伝的性質。形質は量的形質と質的形質に分けられ、前者は体長、体重、肉質、大きさなどの連続的な形質であるのに対し、後者は本数、紋様の数、それらの有無、色やヒトのABO式血液型など不連続な形質。 |
||
ハーディー=ワインベルグの法則 Hardy-Weinberg’s law理想集団において、アリル (遺伝子) 頻度は世代が変わっても変化しないという法則。しかし、現実には有効な集団のサイズは有限であるため、アリル頻度は遺伝的浮動によって変動する。ここでいう理想集団とは以下の通り。1) 集団サイズが非常に多く、2) 他集団の流入・他集団への流出がないこと。3) 有性生殖し、4) 集団内で任意交配 (自由交配) が行われていること。5) (突然) 変異が起きず、6) 特定の遺伝型や表現型に対して自然選択が起きていないこと。 |
||
有効集団サイズ (NE) effective population size自然環境中 (自然の海・川・山) に生息する生物の個体数、もしくは養殖場での飼育個体数は「見かけの大きさ」と言われる量であり、これらすべてが繁殖に関与できるわけではない。ある集団において、実際に次世代へ遺伝的影響を及ぼす個体数の数のことを有効集団サイズと呼ぶ。また、生物集団を存続させるのに必要最小限の集団サイズのことを極限生存可能集団 (minimum viable population; MVP) と呼ぶ。 |
||
人工種苗 artificial seeds養殖または漁獲された親から人間の管理下で (人工的に) 生産された幼生・幼体や稚魚のこと。これに対して、自然環境中で採集した幼体や稚魚を天然種苗natural seedsと呼ぶ。カキ類やアコヤガイの天然種苗は、海中に垂下した採苗基質 (例; ホタテ殻や杉葉) に稚貝を付着させて得る。人工種苗は自然環境に依存する天然種苗と比較して、計画的かつ安定した種苗生産が行え、系統の作出に関連するブランド産物の開発、および食の安全を確保するトレーサビリーティーなどの多くのメリットが挙げられる。また、天然資源保護を目的とした、天然海域への人工種苗放流 (栽培漁業) も行われている。しかし、選抜育種され遺伝的に明らかに異なる系統、あるいは少ない数の親 (自然集団に由来する親であっても) から作出された人工種苗を大量に放流すると、自然集団の遺伝的撹乱を引き起こす要因となるため、これらを考慮した栽培漁業が必要。 |
||
アコヤガイ pearl oyster二枚貝類の一種で、真珠をつくる母貝として知られる。 |
||
アマノリ日本人になじみの深い食用海藻であるノリ(海苔)は,紅藻ウシケノリ目ウシケノリ科アマノリ属(Pyropia)に属する種類である。アマノリ属は世界で130種以上,日本からは29種の生育が報告されており,多くの国で食用として天然物が採集されるとともに,日本,韓国,中国などでは大規模な養殖も行われており,重要な水産物である。生態 アマノリは配偶体である葉状体と胞子体である糸状体とが交互に世代交代を繰り返す生活史をもっている。海苔として食用とされる葉状体は数~数十cmの大きさになる一方,糸状体は顕微鏡的な大きさで,太さ5µm程度のよく枝分かれする体である。葉状体は潮間帯から漸深帯の岩や構造物,他の海藻などに着生するのに対して,糸状体は貝殻などの石灰質の物に穿孔して生育している。 |
||
アオサ・アオノリ類アオサ類が2層膜状,アオノリ類が1層管状,ヒトエグサ類が1層膜状と形態的に区別できるが,市場ではヒトエグサを「アオサ」と呼んだり,「青のり」と明記されていたりする。沖縄の「アーサー」はヒトエグサのことである。以前はアオサ属(Ulva)とアオノリ属(Enteromorpha)は分類学的に区別されていたが,分子系統学的解析が進みアオサ類とアマノリ類は系統的に混ざり合うことが明らかになり,現在ではアオサ類とアマノリ類はまとめてアオサ属に所属している。アオサ類は世界で約100種,日本では20種が報告されている。ヒトエグサ類にはヒトエグサ属(Monostroma)とシワヒトエグサ属(Protomonostroma)があり,ヒトエグサ属が世界で27種,日本で7種が報告されている。 |
||
キチン・キトサン chitin/chithosanキチンは甲殻類の殻などに含まれる多糖類で、キチンをアルカリ処理するとアセチル基が取り除かれキトサンとなる。自然治癒力を高める効果がある。 | ||
バイオミミクリー biomimicry生物の優れた生理機能、形態などを模倣し工学、医学分野などに応用すること。 | ||
採苗 seed養殖、放流のために有用水産動物の卵、幼生や有用海藻類の胞子を野外もしくは養殖場から採取すること。 |
養殖のためホタテガイにカキを付着させる | |
三倍体 triploid本来倍数であるはずの染色体を奇数である3対に人為的に操作したもの。減数分裂がうまくいかず成熟しないが、体細胞は大型化するので、カキ養殖などでは薬品処理によって三倍体を作成して身入りをよくしている。 | ||
極体 polar body卵母細胞が減数分裂をして卵子ができる過程で形成される3つの小さな細胞。核はあるが、細胞質をほとんど持たない。 | ||
付着生物 sessile organism水中基盤に付着して生活する生物。フジツボ類、イガイ類、ゴカイ類、カイメン類、海藻類なども含む。船底、海洋建築物、漁網、食用海藻類に付着する場合は汚損動物と呼ばれる。 |
汚損動物 | |
マボヤ Halocynthia roretzi脊索動物門ホヤ綱に属する一種で北日本、韓国では食され、養殖もされている。[参考 脊策動物門関連図] | ||
生理活性物質 bioactive substance生体に作用し、さまざまな生体反応を制御する化学物質。海綿動物やコケムシ動物から分離されることが多いが、実際には共生細菌類が生産していると考えられている。 | ||
八方サンゴ類 octocoral刺胞動物門花虫綱の1つの分類群で、羽状突起のある8本の触手を備える。宝石サンゴはこの分類群に属する。[参考 刺胞動物門関連図] | ||
テトロドトキシン、TTX tetrodotoxinフグ毒の本体である神経毒で、神経や骨格筋の細胞膜のナトリウムチャンネルに特異的に結合、細胞外から細胞内へのナトリウムイオンの流入を阻止することで細胞膜上の興奮伝達を停止させる。フグ以外でも、Atelopus属のカエル、オウギガニ類、モミジガイ類、ヒョウモンダコ、キンシバイ、ヒモムシ類等、多様な生物相から存在が確認されている。 | ||
テングサテングサ(天草)は,文字通り「寒天をとる草」であるが,テングサという標準和名の海藻はなく,代表種のマクサGelidium elegansやテングサ科藻類(国産27種)のことをいう。地域によっては,同じテングサ属のオオブサ,オニクサ,キヌクサのほか,オバクサ属のオバクサ,ヒラクサ属のヒラクサなども採取されるが,これらの種はマクサと比べて漁獲量も少なく,生態学的な知見も乏しい。 |
||
ミオグロビン mioglobin鉄を含む赤色のポルフィリン色素。魚類の筋肉中に含まれ、この含有量によって赤身魚か白身魚かを区分している。ミオグロビンの主な役割は酸素と結合し運動時に必要な酸素を筋肉内に蓄えることである。 | ||
カロテノイド carotenoid多くの動植物に分布する黄・橙・赤系統の脂溶性色素で、主成分をアスタキサンチン(鮮赤色の色素)とする。ニンジンやトマト、卵黄、フラミンゴなどの着色の他、水産物ではサケ・マスの筋肉色素、魚皮(マダイ等の赤魚)、甲殻類(エビ・カニ)等に含まれる。 | ||
K値 K‐value魚肉の新鮮さに焦点を合わせた鮮度指標であり、鮮度の低下と共に値は大きくなる。魚類の死後変化の過程でATPが急激に失われ、IMP(イノシン酸)が蓄積することから、ATP関連物質中のHX(ヒポキサンチン)とHXR(イノシン)の割合「(HX+HXR/ATP+ADP+AMP+IMP+HXR+HX)×100」で求められる。 | ||
バナースペース

.
本サイトに関するお問い合わせは下記のメールアドレスよりお願いします。
e-mail:takeemon○fishlab.hiroshima-u.ac.jp ○を@に変えてください